")
")



Yuan-Cheng Fung (1919-2019) & Shu Chien (1931-)
Préambule
La biomécanique cellulaire est une discipline récente qui s’est développée à l’initiative de plusieurs chercheurs mais il n’est pas facile de citer un seul pionnier dans cette aventure.
En remontant aux années 60, on ne peut qu’être impressionné par les premiers travaux de S. Chien (92 ans) sur la viscosité du sang [1-3], qu’il relie à la forme et aux propriétés mécaniques des cellules individuelles, i.e. les globules rouges. Mais on ne ne saurait aborder ce thème sans parler du travail essentiel de Y. C. Fung (décédé à l’âge de 100 ans en 2019) qui a travaillé de longues années à faire le lien entre l’échelle cellulaire et l’échelle tissulaire, ceci illustré dans de nombreux ouvrages [4-5] et travaux de recherche.
Plus récemment, E. Evans, D. Ingber ou J. Fredberg ont apporté des contributions importantes aussi bien portant sur la caractérisation que sur la modélisation en biomécanique cellulaire.
Introduction
Shu Chien est né à Beijing en 1931. Sa famille est de descendance royale et la plupart de ses parents ont eu des carrières importantes. Après avoir étudié à Beijing et Taïwan, S. Chien obtient un PhD en 1957 à Columbia University et un doctorat de médecine du NTU [6]. Il publie trois papiers fondamentaux dans la revue Science sur la viscosité du sang. Ce travail est très original, car il explique les propriétés macroscopiques du sang (viscosité), avec un éclairage physique nouveau reposant sur la déformabilité des globules rouges (GRs), qui s’allongent, adhèrent et forment des rouleaux (agrégation) en fonction du gradient de cisaillement.
En particulier, S. Chien propose une expérience originale pour mesurer la capacité des GRs [1,2] à se déformer au travers de capillaires (Fig. 1, tailles ~ 2-7µm, plus petits que le diamètre des érythrocytes ~ 8µm) : c’est une première en microfluidique ! En outre, S. Chien et ses collègues proposent une explication microscopique des effets non-Newtoniens (de viscosité) observés, en lien avec l’adhérence, l’agrégation et l’effet de la concentration en fibrinogène [3]. En définitive, ce travail permet de montrer comment les propriétés à l’échelle microscopique peuvent se révéler essentielles pour la compréhension de phénomènes aux échelles supérieures.
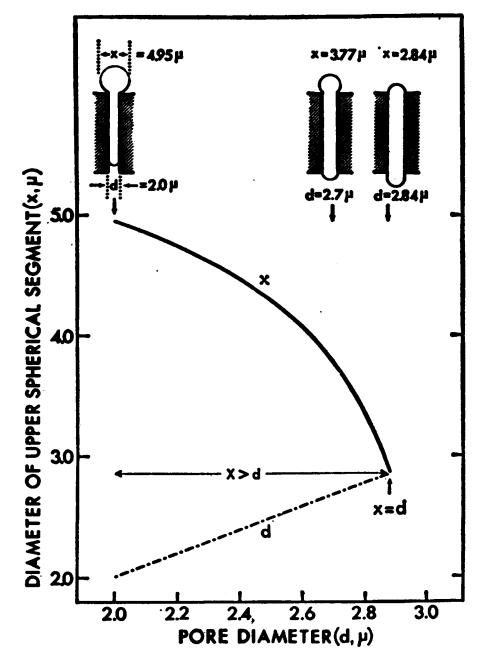
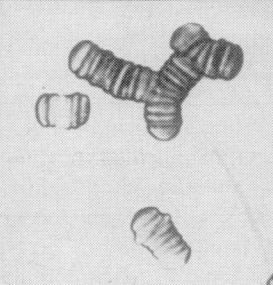
Figure 1. A gauche, filtration de GRs au travers de tamis de diamètres différents [1] et évolution de la dimension amont du GR en fonction du diamètre du pore (µm). A droite, agrégation de GRs avec formation de rouleaux [3].
Par la suite S. Chien sera nommé Professeur de physiologie et biophysique à l’Université de Columbia. Il devient ensuite Professeur à l’Université de Californie à San Diego (UCSD) où il dirigera le département de Bioingénierie jusqu’en 2005. Il a reçu de nombreuses récompenses au cours de sa carrière (Chinese Academy, Academy of Sciences, USA, National Medal Of Science, The White House, Benjamin Franklin Medal in Mechanical Engineering, etc.)
Yuan-Cheng Fung est aussi un précurseur dans le domaine des approches biomécaniques pour le vivant. Né en 1919, il étudie à l’Université de Chine jusqu’en 1943. Puis il émigre aux Etats Unis où il obtient un PhD à Caltech en 1948 en aéronautique et mathématiques. Sa première carrière est en aéroélasticité, domaine auquel il se consacrera pendant 20 ans !
Pendant cette période, il écrit des ouvrages en mécanique des solides et des milieux continus. En 1958, il décide de se consacrer à la biomécanique qui est alors une science nouvelle. Quelques années plus tard, il initie en 1966 un programme en bioingénierie à l’université de Californie à San Diego (UCSD). Il se consacre alors à la circulation sanguine, et à la biomécanique des tissus vivants. Il est le précurseur de la modélisation rhéologique des globules rouges d’un point de vue continu, et il a notamment proposé des modélisations pertinentes du globule rouge en prenant en compte à la fois la membrane et la partie interne de la cellule, i.e. le cytoplasme (Fig. 2).
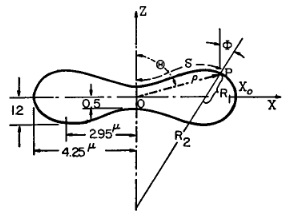
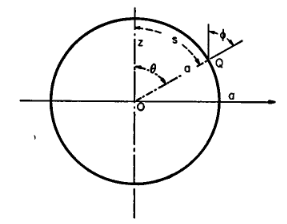
Figure 2. Modélisation du globule rouge, un fluide visqueux entouré d’une membrane 2D élastique dont la rigidité varie le long de la surface [7].
Dans les années 80, Y.-C. Fung a rédigé de nombreux ouvrages pionniers du domaine, qui sont aujourd’hui considérés comme des références [5-6], aussi bien par les biomécaniciens et physiciens que par les biologistes. Il part des propriétés microscopiques pour aller vers le continu, domaine de la mécanique qu’il maîtrise parfaitement. Dans la période plus récente, il s’est consacré à la compréhension des lois de croissance tissulaire et du processus de vieillissement.
Il a reçu de nombreuses récompenses ou a été élu dans des sociétés prestigieuses (Caltech, Jap. Society of Mechanical Eng., National Acad. Eng., Institute of Medicine 1991, Natl. Acad. Sci, 1992, Chinese Acad. Sci., Academia Sinica, Taipe).
Si les travaux de S. Chien et Y.-C. Fung restent assez simples de part leurs applications aux GRs, il n’en reste pas moins qu’ils ont apporté des réponses essentielles en se plaçant à des échelles microniques, afin de mieux appréhender la mécanique à l’échelle cellulaire. Le concept de biomécanique cellulaire s’est ainsi développé constamment depuis cette époque, et au cours des 50 dernières années ; cela a aussi révélé que les approches multi-échelles - certes difficiles à mettre en œuvre - sont les seules qui permettent de faire émerger des mécanismes de fonctionnement biologiques. Mais voyons tout d’abord comment ces travaux pionniers ont permis d’améliorer la compréhension du vivant et de stimuler de nouvelles approches.
Des globules rouges aux cellules eucaryotes
Les premiers travaux, comme indiqués précédemment, ont beaucoup porté sur le globule rouge, qui est l’objet cellulaire le plus facile a étudier. Cependant les cellules eucaryotes ne sont pas uniquement dotées d’une membrane entourant un fluide ; elles possèdent un noyau, un cytoplasme, des organites et tout un réseau de filaments interconnectés appelé cytosquelette. En outre, certaines protéines du cytoplasme sont capables de se lier à des protéines transmembranaires pour créer des sites adhésifs permettant aux cellules de se déplacer et/ou d’adhérer à d’autres cellules ou substrats.
Néanmoins de nombreux travaux sont restés, un temps, focalisés sur les GRs, ce qui a permis de développer de nouvelles méthodes de mesures mécaniques. E. Evans a proposé d’utiliser des micropipettes [8] afin d’aspirer des GRs au travers d’une pipette au sein de laquelle la pression est contrôlée. Ainsi, des modèles basés sur les études de Y.-C. Fung ont pu être développés permettant d’améliorer le modèle du GR, par exemple en tenant compte du cytosquelette de spectrine sous-jacent. Ces études se sont développées ensuite dans le cadre d’autres cellules vivantes possédant un noyau, comme les globules blancs. Dans ce contexte, les modèles de capsules [9-10] ou vésicules, ainsi que le modèle de goutte composite (une enveloppe, la membrane entourant un cytoplasme visqueux et un noyau visqueux ou élastique) ont été proposés, afin d’améliorer les approches précédentes.
Dans les années 90, D.E. Ingber, à partir d’idées originales venant des structures en génie civil, propose de modéliser la cellule grâce au modèle de tenségrité, qui prend en compte les efforts internes de traction et de compression [11]. Cette étude sera importante car elle va permettre des développements en lien avec les nouvelles techniques émergentes de microscopie de fluorescence qui donnent accès aux divers filaments du cytosquelette (filaments d’actine, microtubules, filaments intermédiaires). C’est une illustration du fait que le changement d’échelle doit s’appuyer aussi sur ce qui se passe au niveau des nanostructures cellulaires. On descend encore d’une échelle.
Grâce aux différents modèles explicités ci-dessus, après ajustement expérimental des données mécaniques, on va pouvoir prédire la déformabilité des cellules en conditions physio-pathologiques : circulation, roulement des cellules circulantes tels les globules blancs ou les cellules tumorales [12], près de la paroi vasculaire, interactions cellule-paroi, transmigration, migration sur substrat 2D ou 3D dans un réseau de fibres, etc.
La biomécanique cellulaire de nos jours
Bien sûr, les modèles décrivant les cellules ne sont pas parfaits, c’est pourquoi de nouvelles techniques instrumentales sont maintenant de plus en plus développées pour appréhender le comportement complexe cellulaire. Comme vu précédemment, certains éléments de la cellule ont un comportement élastique, visqueux, voire viscoélastique.
Dans cette optique, des expériences originales modernes ont été proposées : pinces optiques, pinces magnétiques, microfluidique, Microscopie à Force Atomique (AFM), microplaques, suivi de particules intracellulaire, pour les mesures locales sur cellules isolées [13]. Une des questions qui se pose est de savoir s’il faut tester une cellule isolée (en suspension dans un fluide) ou adhérente, ce qui donne parfois un comportement très différent. De même, les propriétés mécaniques cellulaires dépendent de l’environnement car les cellules sont mécano-sensibles, il est donc primordial de tester des cellules sur (ou dans) un substrat proche de l’environnement réel : en effet, des myocytes ne réagissent pas de la même manière que des neurones au contact d’un substrat [14].
Dans ces expériences de mécanique cellulaire, une autre question importante qui se pose est de savoir quel est le niveau de contrainte à appliquer sur la cellule [15]. Ce niveau de contrainte/force dépend à la fois de la technique utilisée mais aussi du choix de la sonde mécanique utilisée, de sa taille et de sa rigidité. Le protocole expérimental de chargement mécanique est également très important.
Au niveau des mesures globales, les chambres d’écoulement, la rhéométrie, la magnétocytométrie, les systèmes d’étirement de substrat sont des méthodes qui portent sur le comportement d’un ensemble de cellules, permettant ensuite de moyenner les comportements, ce qui est utile pour obtenir des données statistiques significatives.
Grâce à ces nouveaux outils, la mesure de la déformabilité proposée par S. Chien devient de plus en plus précise et les comportements étudiés (viscoélasticité, poroélasticité) deviennent accessibles pour comprendre les phénomènes physio-pathologiques, en particulier la migration cellulaire. Dans le cancer en particulier, la diminution des propriétés élastiques va de pair avec la malignité ou l’invasivité des cellules [16]. Outre ces études, le cytosquelette de la cellule est devenu le centre d'intérêt avec le développement de concepts visant l'activité cellulaire (acto-myosine), la stabilité des filaments cellulaires, la dynamique cellulaire, et J.J. Fredberg a proposé le concept de rhéologie vitreuse qui donne lieu à des comportements rhéologiques en loi puissance [17]. Les études ont largement porté sur les cellules en adhérence en 2D, et maintenant en 3D. La mécano-transduction (conversion d'un signal mécanique en signal chimique et inversement) a aussi été mise en évidence.
Quelques méthodes originales récentes en biomécanique cellulaire sont :
- la microscopie à force de traction (TFM) qui permet de mesurer les forces exercées par les cellules sur leur environnement [18], tel un substrat ou un milieu fibreux 3D. Elle trouve notamment beaucoup d’intérêt pour la compréhension des mécanismes en jeu dans la migration des fibroblastes ou des cellules musculaires [19].
- la mesure de l'adhésion cellule-cellule [20] et cellule-substrat par AFM et micropipettes. C'était aussi l'une des observations de Chien en 1967 qui a permis la compréhension du mécanisme de formation des rouleaux.
- la migration individuelle [21] ou collective [22] de cellules, utilisant les techniques de PIV (Vélocimétrie par Imagerie de Particules) modernes dans le contexte des déformations d’une monocouche de cellules (épithéliales par exemple).
- la compréhension des forces issues de mécanismes actifs, au niveau des cellules musculaires [23] et dans la dynamique de l’acto-myosine au cours de la migration cellulaire [24].
Ce sont donc ces nouvelles approches, qui aujourd’hui permettent d’étudier le comportement de la cellule dans un environnement complexe, grâce à des modèles biomécaniques, prenant en compte à la fois la complexité interne de la cellule, mais aussi les couplages avec les interfaces (matrice extra-cellulaire ou autres cellules).
Les applications aux échelles supérieures semblent maintenant possibles et s'orientent vers les modèles biologiques d'organoïdes, au sein desquels les propriétés mécaniques cellulaires jouent un rôle essentiel. Ici, un des enjeux reste encore de comprendre comment passer d'un système microscopique (la cellule) à un système complexe macroscopique (le tissu), à l’instar de ce qu’a proposé Y.-C. Fung dans ses travaux pionniers.
Conclusion
La biomécanique cellulaire est omniprésente aujourd'hui. Les enjeux ont aussi beaucoup évolué grâce au développement de techniques avancées en microscopie de fluorescence (confocale, STED, etc.) ou rayonnements X, permettant le marquage et la reconnaissance des constituants cellulaires en 3D. Ceci donne des informations microscopiques précieuses permettant d’élaborer ou affiner des lois de comportement basées sur la réalité intracellulaire.
Shu Chien et Yuan-Cheng Fung ont amené un éclairage essentiel qui a permis des avancées significatives, dont la plupart inspire encore les chercheurs en biomécanique cellulaire. Les progrès actuels laissent espérer que les nouveaux outils expérimentaux et de modélisation permettront encore de grands progrès, afin de mieux comprendre la dynamique spatio-temporelle cellulaire.
Références
[1] Gregersen, M.I., Bryant, C.A., Hammerle, W. E., Usami, S. & Chien, S. Flow Characteristics of Human Erythrocytes through Polycarbonate Sieves, Science, 157, 825-827 (1967)
[2] Chien, S., Usami, S., Dellenback, R.J. & Gregersen, M.I. Blood viscosity: influence of erythrocyte deformation, Science, 157, 827-829 (1967)
[3] Chien, S., Usami, S., Dellenback, R.J., Gregersen, M.I., Nanninga, L.B. & Mason-Guest, M., Blood viscosity: influence of erythrocyte aggregation, Science, 157, 829-831 (1967)
[4] Fung Y.-C., Biomechanics. Mechanical properties of living tissues, 2nd Edition, Springer (1993)
[5] Fung Y.-C., Biomechanics. Circulation, 2nd Edition, Springer (1996)
[6] https://en.wikipedia.org/wiki/Shu_Chien
[7] Fung Y.-C. & Tong. P., Theory of the sphering of red blood cells, Biophys. J. 8, 175-198 (1968)
[8] Evans, E. & Yeung, A., Apparent viscosity and cortical tension of blood granulocytes determined by micropipette aspiration, Biophys. J., 56, 151-160 (1989)
[9] Secomb, T.W., Skalak, R., Ozkaya, N. & Gross, J.-F., Flow of axisymmetric red blood cells in narrow capillaries, J. Fluid Mech., 163, 405-423 (1986)
[10] Barthès-Biesel, D. & Rallison, J.M. The time-dependent deformation of a capsule freely suspended in a linear shear flow, J. Fluid Mech., 113, 251-267 (1981)
[11] Ingber, D.E., Cellular tensigrity: defining new rules of biological design that govern the cytoskeleton, J. Cell Sci. 104, 613-627 (1993)
[12] Dembo, M., Torney, D.C., Saxman, K.D.
The reaction-limited kinetics of membrane-to-surface adhesion and detachment, Proc. R. Soc. Lond. B, 234, 55-83 (1988)
[13] Verdier, C., Rheological properties of living materials. From cells to tissues. J. Theor. Medicine, 5, 67-91 (2003)
[14] Discher, D.E., Janmey, P., Wang, Y.-L., Tissue cells feel and respond to the stiffness of their substrate. Science, 310, 1139-1143 (2005)
[15] Laurent V.M., Hénon S., Planus E., Fodil R., Balland M., Isabey D. & Gallet F., Assessment of mechanical properties of adherent living cells by bead micromanipulation: comparison of magnetic twisting cytometry vs optical tweezers. J Biomech Eng., 124, 408-421 (2002)
[16] Lekka, M., Laidler, P., Gil, D., Lekki, J., Stachura, Z. & Hrynkiewicz, A.Z. Elasticity of normal and cancerous human bladder cells studied by scanning force microscopy. Eur. Biophys. J. 28, 312-316 (1999).
[17] Bursac, P., Lenormand, G., Fabry, B., Oliver, M., Weitz, D.A., Viasnoff, V., Butler, J.P. & Fredberg J.J., Cytoskeletal remodelling and slow dynamics in the living cell. Nat. Mater., 4, 557-561 (2005)
[18] Dembo, M., Wang Y. L. Stresses at the cell-to-substrate interface during locomotion of fibroblasts. Biophys J., 76, 2307-2316 (1999)
[19] Tracqui, P., Ohayon, J. & Boudou, T.
Theoretical analysis of the adaptive contractile behaviour of a single cardiomyocyte cultured on elastic substrates with varying stiffness.
J. Theor. Biol., 255, 92-105 (2008)
[20] Benoit, M., Gabriel, D., Gerisch, G. & Gaub, H.E. Discrete interactions in cell adhesion measured by single-molecule force spectroscopy. Nat. Cell Biol., 2, 313-317 (2000)
[21] Di Milla, P.A., Barbee, K. & Lauffenburger, D.A. Mathematical model for the effects of adhesion and mechanics on cell speed. Biophys. J., 60, 15-37 (1991)
[22] Petitjean, L., Reffay, M., Grasland-Mongrain, E., Poujade, M., Ladoux, B., Buguin, A. & Silberzan P. Velocity fields in a collectively migrating epithelium. Biophys. J., 98, 1790-1800 (2010)
[23] Harris, D.E. & Warshaw, D.M., Length vs. Active force relationship in single isolated smooth muscle cells, Am. J. Physiol., 29, C1104-C1112 (1991)
[24] Pollard, T.D., Borisy, G.G. Cellular motility driven by assembly and disassembly of actin filaments. Cell, 112, 453-465 (2003)
Claude VERDIER* (DR CNRS)
Valérie M. LAURENT* (MCF UGA)
*Equipe Migration des Cellules en Milieu Complexe (MC2), Laboratoire Interdisciplinaire de Physique, UMR 5588, 140 rue de la physique, 38400 Saint Martin d’Hères